|
|
|
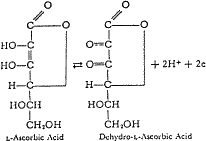
Figure 1. The Reversal Oxidation-Reduction |
The Vitamin C Conspiracy Articles proving Vitamin C cures infections
Irwin Stone is a chemical engineer by training, a biochemist by vocation and a paleopathologist by avocation. He started working on ascorbic acid in 1934 and received the first U.S. Patents for its use as a food antioxidant. Worldwide use of this process has been a factor in the virtual elimination of frank clinical scurvy in the developed countries. In 1965-67 he published a series of papers describing the genetic, liver-enzyme disease, Hypoascorbemia and its significance in Medicine. This new genetic approach supplies the rationale for the use of massive doses of ascorbic acid in Orthomolecular Therapy. He is now Research Director of Megascorbic Research, Inc.
Ascorbic acid is a vital, ubiquitous substance, in the life process. All living organisms either make it, get it in their foodstuffs or they perish. The enzyme systems for the production of ascorbic acid are of ancient origin and were formed very early in the development of the life process on this planet, probably while the most highly developed forms were still primitive unicellular forms.
The evidence of both plant and animal embryology corroborates this viewpoint as the dormant plant seed and animal egg are devoid of ascorbic acid. There is an immediate production of ascorbic acid in the germinating seed or developing egg, even when the embryo is nothing more than a cluster of a few cells.
Its widespread occurrence in all present day multicellular organisms, both plant and animal, also testify to this. We can also infer that ascorbic acid production was well developed before the living organisms diverged into the plant and animal forms, Stone.1
Chemically, ascorbic acid is a simple carbohydrate material, related to glucose, of rather unique properties. The presence of the ene-diol group in the molecule confers electron lability, which makes it a member of an oxidation-reduction system having electron donating and electron accepting properties, see Fig. 1. At a submolecular level, the living process is nothing more than a stepwise orderly, transfer of electrons, so that the presence of an electron-labile system such as ascorbic acid in a living organism acting in concert with other ancient oxidative-reductive systems, aids in maintaining electron-transfer efficiency in the living process.
|
|
|
Figure 1. The Reversal Oxidation-Reduction |
If we rely on the examination of present day living animals for their ascorbic acid production to estimate its occurrence in the fossil record, we accumulate much interesting and enlightening data on the evolution of the enzyme systems involved. Based on the known accuracy of the genetic transfer of information, we can assume that the enzyme systems and their bodily location in present living animals have changed little from their ancient representatives.
Every one of the primitive invertebrates and lower organisms so far examined showed the presence of ascorbic acid, Bourne and Allen;2 Bourne.3 In nearly all the vertebrates examined, ascorbic acid production is the normal state. Those few species that cannot make their own ascorbic acid are suffering from a genetic defect in their enzyme-production systems and must receive a supply of ascorbic acid from their foodstuffs or die of scurvy.
Table 1 shows the locus of the ascorbic and enzyme system during the course of evolution of the vertebrates from the fishes to the primates, based on data obtained by the examination of present day animal forms, Chatterjee et al.1 Roy and Guha5; Chaudhuri and Chatterjee.6
The locus of the enzymes for ascorbic acid production in the cold-blooded vertebrates, the fishes, the amphibians and the reptiles, are in the kidneys. The more highly active warm-blooded mammals all synthesize their ascorbic acid in their liver. One of the main functions of ascorbic acid in animal physiology is the maintenance of biochemical homeostasis under stress. The greater the stress an animal undergoes, the more ascorbic acid it produces.
About 165 million years ago, when Nature had the evolution of the more active and stressful mammals in view, an important morphological and physiological decision had to be made. The kidneys, while adequate as the site of ascorbic acid synthesis for the rather sluggish cold-blooded vertebrates, were inadequate for the increased ascorbic acid needs of the more highly stressed mammals. The successful solution of this problem was the transfer of the enzymes for the production of ascorbic acid from the relatively small biochemically-crowded kidney to the more spacious liver, which is the largest organ of the body. All present day mammals capable of synthesizing ascorbic acid are liver producers because any ancient form which did not make this transfer was so biochemically handicapped that they were eliminated by the forces of Evolution.
The present day birds, whose ancestors appeared about the same time as the mammals, still show this kidney-liver transition, Chaudhuri and Chatterjee.6 The older order of present day birds, such as the ducks, pigeons and hawks, synthesize their ascorbic acid in their kidneys, while in the more recent order of the perching and song birds, the Passeriformes, some produce ascorbic acid both in their kidneys and livers, others only in their liver. Some, like man, are incapable of synthesizing ascorbic acid at all.
As Evolution proceeded, the primates appeared about 65 million years ago and like other mammals should be capable of synthesizing ascorbic acid in their livers. However, something happened during the evolution of the primates because it has been known for thousands of years that Man, unlike other mammals, was susceptible to scurvy.
Up until 1907 scurvy was considered a completely human disease as no other animal was known to be susceptible to it. In 1907, Holst and Frohlich7, working on ship beri-beri contracted aboard ship for the Norwegian Fishing Fleet, wanted a small mammal to substitute as a test animal for the pigeons then used. They fed guinea pigs the test diet, which produced beriberi in their pigeons, and much to their surprise, scurvy resulted instead. Later it was shown that laboratory monkeys were also susceptible to scurvy. Man, guinea pigs and certain monkeys, unlike other mammals, cannot make their own ascorbic acid.
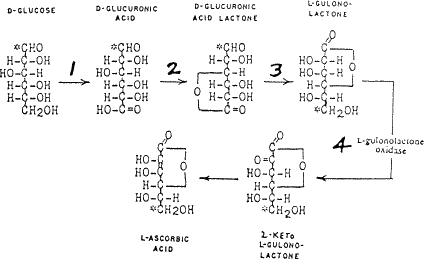
In 1912 the vitamin hypothesis was postulated (Funk8), part of which stated that scurvy was a deficiency disease caused by a lack of an unknown water-soluble substance, called Vitamin C, in the diet. Twenty ears later in 1932 (Svirbely and Szent-Györgyi ), it was demonstrated that ascorbic acid was identical with Vitamin C. Burns10 in 1959 Showed that the basic biochemical lesion in the few mammals susceptible to scurvy was due to their inability to produce the active enzyme, L-gulonolactone oxidase, involved in the in the mammalian conversion of blood glucose to ascorbic acid, in their livers. This synthesis, involving four enzymes, is illustrated in Fig 2. Man has the first three enzymes in his liver but it is the missing fourth enzyme, which completely blocks the liver production of ascorbic acid.
|
|
|
Figure 2. Man has the first three enzymes in his
liver but it is the missing fourth |
Up until 1965 it was assumed that all primates were unable to produce their own ascorbic acid and were thus susceptible to the disease, scurvy. It was pointed out (Stone1) that this was merely an assumption and one which had never been tested. It was suggested that the whole order of the primates should be examined for the presence of L-gulonolactone oxidase in their livers. If this was done, then the data obtained might be useful in pinpointing, in time, when the mutation occurred. Thus it might be possible to determine in which primate ancestor of man this important enzyme system was lost.
This suggestion was picked up and tests were reported from Harvard (Elliott et al.11) in 1966 and Yerkes Primate Research Center in 1969 (Nakajima et al.12) wherein it was indicated that all of the monkeys examined, which were members of the suborder, Prosimii, showed active enzyme, L-gulonolactone oxidase, in their livers while the livers of those members of the suborder Anthropoidea are inactive. While the data is not fully complete for all members of the primate order, the present sampling indicates that the dividing line between those primates which are susceptible to scurvy and those that arc not, is likely to be between the two main suborders, the Prosimii and the Anthropoidea.
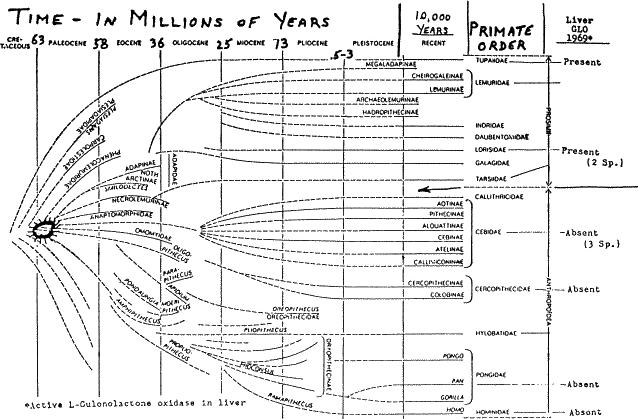
This is illustrated in Fig. 3 showing a chart of the fossil record of the primates as devised by Simons.13 The second column from the right is a listing of the families of the present day primates. The column on the far right, marked “Liver GLO” contains the results of the recent examination of the primate livers for the presence of the active enzyme, L-gulonolactone oxidase. In all the primates thus far examined, those in the six genera of the suborder, Anthropoidea, lack this enzyme in their liver while those of the four genera Prosimii have the active enzyme and can synthesize their own ascorbic acid. While more members of other Prosimii families should be examined such as Lemuridae, Tarsiidae and Daubentoniidae, the present indications are that the division between the Prosimii and the Anthropoidea is not only morphological but also in this important physiological parameter.
|
|
|
Figure 3. Chart of the Fossil Record of the
Primates with the Occurrence of Active L-Gulonolactone Oxidase |
If we follow the arrow and extrapolate this dividing line back into time on this chart, then we arrive at a point between the late Cretaceous and late Paleocene where this mutation which destroyed the gene for the synthesis of the enzyme protein, L-gulonolactone oxidase, appears to have occurred. This happened not long after the primates appeared on the scene, about 58 to 63 million years ago in the vicinity of the spurred circle, in the primate ancestor whose progeny evolved into the suborder Anthropoidea.
It is probably more than coincidental, that in the neighborhood of the late Cretaceous period, when this primate genetic accident occurred, there was a brief interval when many other organisms underwent a severe attenuation in diversity or became extinct. This included many invertebrate forms and notably the vertebrate dinosaurs which suddenly disappeared from the fossil record. Russell and Tucker14 in 1971 suggested that a nearby supernovae explosion, with its liberation and absorption by the earth’s atmosphere of large fluxes of cosmic rays gamma rays and x-rays, might have produced climatic effects so drastic as to have caused the extinction of many animals including the cold-blooded dinosaurs. If such an astronomical event occurred, then the random absorption of some of this high energy radiation may have been instrumental in mutating the primate gene for the synthesis of the enzyme protein, L-gulonolactone oxidase, resulting in an inactive enzyme.
Thus a conditional lethal mutation (Gluecksohn-Waelsch15) happened to this primitive primate. The destruction of so vital a biochemical process would have had lethal consequences were it not for the fact that it occurred to an arboreal animal living in a tropical or semi-tropical environment where plenty of foodstuffs containing ascorbic acid were available throughout the entire year. The diet of the mutated primate may not have supplied as much ascorbic acid as its previous liver synthesis, but it was sufficient for survival. Bourne16 in 1944 showed that a modern gorilla, living in its natural habitat, would obtain 4.5 grams of ascorbic acid per day from its foodstuffs.
Pauling17 in 1970, basing his calculations on the caloric content and ascorbic acid levels in raw plant foods, concluded that the range of the optimum intake is about 2.3 to 9.5 grams per day. He also pointed out, while the range of the B vitamins in 110 raw plant foods supplying 2,500 calories was only two to four times the recommended dietary allowances, the corresponding ratio for ascorbic acid was 35 times that recommended, 2,300 milligrams versus 60 milligrams a day.
Pauling indicated in 1968 that this mutation may have had survival value at the time because it freed the biochemical machinery for other purposes and conserved energy. This survival value was lost as soon as the progeny of this mutated animal, evolving into the future genus, Homo, left the trees, moved into temperate climes and changed its diet to one where high levels of ascorbic acid were not dominate the year round.
Man still carries this defective gene and it has no survival value for modern Man. In fact, in the course of prehistory and during historical times, it has been a severe handicap and the side effects of this defective gene have resulted in the deaths of more individuals, caused more sickness and suffering and have changed the course of history more than any other single factor.
As a result of these evolutionary studies, it was indicated by Stone19 in 1966 that due to this defective gene, which produced an inactive enzyme, present day humans are suffering from a mammalian inborn error of carbohydrate metabolism which was named, Hypoascorbemia Scurvy is actually the sequelae of this genetic liver-enzyme disease which makes it necessary for man to obtain ascorbic acid from exogenous sources, Stone.20 For the past 60 years, ascorbic acid has been regarded as “vitamin C” when actually it is a liver metabolite and certainly not a “vitamin” for the myriad of mammals having the intact gene for L-gulonolactone oxidase. These mammals never contract scurvy, even on a diet completely free of vitamin C.
If correction of this inborn error of carbohydrate metabolism is defined as a need to supply, to the individual, the daily amount of ascorbic acid the human liver would be producing, if the intact gene were present, then the daily production of this liver metabolite would be about two to four grams, based on data for the unstressed rat (Burns et al.21 Saloman and Stubbs22) and at least 15 grams per day under stress, Conney et al.23
When this indicated daily multigram synthesis of the “liver metabolite,” ascorbic acid, is compared with the daily amount of “vitamin C,” 60 milligrams (Food and Nutrition Board24), recommended as nutritionally adequate, for humans, there is a 33 to 250 fold disparity. Ever since the vitamin C-dietary disorder hypothesis was postulated in 1912, the emphasis has been on the prevention or cure of the symptoms of frank clinical scurvy to the neglect of the many other basic functions of ascorbic acid in the living process. A subject given the daily few milligrams of vitamin C will not show the classic symptoms of frank clinical scurvy but may still be suffering from severe subclinical biochemical scurvy.
The vitamin C hypothesis which has had unchallenged acceptance since 1912 originated at a time when modern, widely accepted biochemical and genetic concepts were either unknown or unrecognized. The classic lectures of Sir Archibald Garrod25 on the “Inborn Errors of Metabolism,” in which he showed that missing enzymes could cause diseases, were delivered only four years earlier in 1908. This revolutionary medical concept was ignored and neglected for decades. The discovery, isolation and synthesis of ascorbic acid were still twenty years in the future. The modern views on genetic information transfer were unknown. Clearly the 60-year old vitamin C hypothesis with its built-in orientation toward minute doses and the alleviation of acute clinical scurvy is a gross oversimplification of our present knowledge and subordinates the long term effects of chronic inadequate ascorbic acid intakes on the multitude of physiological functions of ascorbic acid in the human body.
In the clinical research of the past 40 years there have been hundreds of papers written to find the daily minimum level of ascorbic acid needed to prevent or eliminate the classic symptoms of frank clinical scurvy. However, you will not find a single paper reporting on the long-term effects of continued administration of ascorbic acid based on time levels produced endogenously in the mammalian liver. These high levels of ascorbic acid production have served so well in maintaining biochemical homeostasis under stress for the past 165 million years during the evolution and world dominance of the mammals. After nearly 60 million years since this primate mutation occurred, it is only in the last 40 years, since the synthetic production of unlimited quantities of ascorbic acid, that it has become possible to fully correct this genetic liver-enzyme disease.
These genetic concepts provide the rationale for the use of large doses (megascorbic levels) of ascorbic acid and open up wide vistas of research in preventive medicine and therapy, Stone,26 In the few areas where megascorbic therapy has been tried, it has been eminently successful; in the treatment of the viral diseases, in glaucoma, in schizophrenia and as a detoxicant. Research on other applications of these genetic principles will lead to the megascorbic prophylaxis and megascorbic therapy of many other disease states, Stone.2
Evidence is presented that time enzyme systems for the important synthesis of ascorbic acid are of very ancient origin, beginning long before time plant and animal lines diverged. The progressive vertebrate evolution of these enzymes are traced through the fishes, amphibians, reptiles, birds and mammals. Shortly after the appearance of the primates, a genetic mutation occurred on the gene for the liver enzyme L-gulonolactone oxidase, which destroyed this animal’s ability to produce ascorbic acid from blood glucose. The progeny of this mutated animal developed in to time present day members of the primate suborder, Anthropoidea. The non-mutated primates are the ancestors of the present suborder, Prosimii.
Extrapolation of this data into the primate fossil record indicates the mutation to have occurred in the period the same nearby supernovae explosion which was possibly responsible for the extinction of the dinosaurs and the disappearance of many invertebrates in the late Cretaceous.
Members of the genus Homo, present day Man, still carry this defective gene and during prehistoric and historical times it has been responsible for more deaths, more sickness and human misery and more changes in history, than any other single factor.
The vital importance of ascorbic acid in many phases of human physiology has been underrated for the past 60 years because in 1912, 20 years before its discovery and synthesis, it was designated a “vitamin” for the treatment of frank clinical scurvy which was considered a simple dietary disturbance.
Actually, ascorbic acid is a liver metabolite produced in nearly all mammals in large daily amounts. Because of this defective gene Man is suffering from a mammalian genetic liver-enzyme disease, a true “inborn error of carbohydrate metabolism,” named Hypoascorbemia. Scurvy is not a distinct disease entity but merely the final fatal sequelae of uncorrected Hypoascorbemia.
This genetic approach provides the rationale for the use of large daily doses of ascorbic acid and opens wide vistas of research for its application to preventive medicine and therapy.
|
TABLE I |
|||
|---|---|---|---|
LOCUS OF ASCORBIC ACID ENZYMES
|
|||
Organism |
Time of Appearance
|
Site of Ascorbic
|
|
| Fishes |
425 |
Kidneys | |
| Amphibians |
325 |
Kidneys | |
| Reptiles |
205 |
Kidneys | |
| Birds |
165 |
||
| Older Orders | Kidneys | ||
| More Recent Orders |
Kidneys and Liver |
||
| Most Recent Orders |
Liver | ||
| Mammals |
165 |
Liver | |
From Orthomolecular Psychiatry, 1972, Volume 1, Numbers 2 & 3, pp. 82-89.
HTML Revised 22 February, 2003.
Corrections and formatting © 2000-2003 AscorbateW